An activated wheat CC G10 -NLR immune receptor forms an octameric resistosome
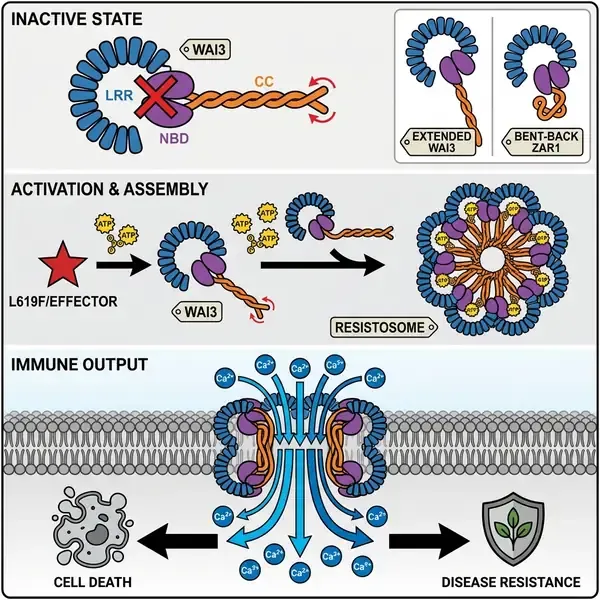
植物核苷酸结合亮氨酸富集重复(NLR)受体的结构研究自五聚体ZAR1抗性小体发现以来经历了范式转移。随后的研究陆续鉴定了四聚体TIR-NLR和六聚体CC-NLR组装体,然而一个重要的系统发育分支——G10型卷曲螺旋(CC_G10)-NLR在结构上一直是个谜。近日,发表在《细胞》(Cell)杂志上的一项合作研究揭示了小麦CC_G10-NLR蛋白WAI3(Wheat Autoimmunity 3)的冷冻电镜结构,发现了一种前所未有的八聚体抗性小体,重新定义了植物先天免疫的结构边界。
该研究始于对小麦自身免疫突变体M3405的鉴定,该突变体表现出自发的超敏反应(HR)样病斑,并增强了对白粉病的抗性。图位克隆将该表型追溯到WAI3亮氨酸富集重复(LRR)结构域中的一个功能获得性(GOF)突变(L619F)。这一单氨基酸替换足以触发组成型免疫激活,为激活态受体的结构表征提供了稳定的平台。
冷冻电镜分析显示,激活的WAI3复合物组装成一个巨大的环状八聚体,直径约为200 Å。这一发现得到了拟南芥RPS2(另一种著名的CC_G10-NLR)在激活后同样形成八聚体的证实,表明这种高阶寡聚化是G10谱系的保守特征。除了化学计量比外,WAI3抗性小体在结构上与此前表征的CC-NLR有显著不同。在ZAR1和Sr35等受体中,CC和LRR结构域通过保守的EDVID基序稳定在核苷酸结合结构域(NBD)的同侧,形成“折叠”构象。然而,WAI3缺乏该基序,导致其呈现出一种“伸展”的结构安排,即CC和LRR结构域位于NBD的相反两侧。这种配置与TIR-NLR抗性小体的架构更为相似,凸显了不同NLR类别之间显著的结构趋同现象。
在功能上,WAI3抗性小体充当钙离子通透通道。在木本烟草中的电生理和钙成像实验表明,WAI3的激活触发了强烈的、多相的胞质钙离子([Ca2+]cyt)内流。这种内流以初始峰值和随后的持续第二相为特征,其动力学模式不同于其他CC-NLR中观察到的瞬时模式。有趣的是,WAI3未能诱导人类HEK293T细胞产生钙电流,表明其通道功能的实现需要植物特有的辅助因子。
此外,研究人员提出了一种基于螺旋结构域1(HD1)与翼状螺旋结构域(WHD)之间夹角来预测NLR寡聚状态的结构指标。他们观察到较小的HD1-WHD夹角与更高阶的寡聚体相关(例如WAI3八聚体为80°,而ZAR1五聚体为92°)。这一“角度准则”为解释AlphaFold建模的NLR结构提供了预测工具。通过填补NLR系统发育图谱中的关键空白,该工作确立了CC_G10-NLR抗性小体作为一类独特的免疫信号枢纽,并强调了植物免疫系统在构建多样化分子支架以防御病原体方面的卓越可塑性。