Deciphering mechanical determinants of morphological evolution
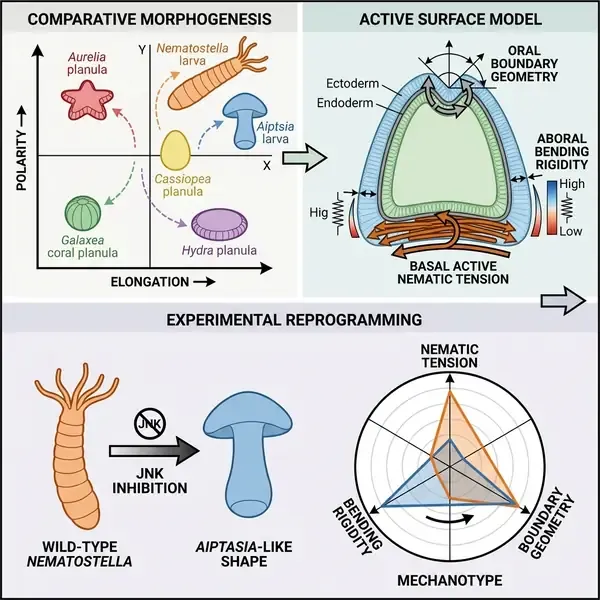
长期以来,演化发育生物学(evo-devo)一直侧重于研究驱动形态多样性的遗传和分子转变。然而,一个根本性的空白仍然存在:这些分子变化是如何转化为实际塑造组织的物理力量的。在发表于《细胞》(Cell)杂志的一项里程碑式研究中,研究人员引入了一个“力学型”(mechanotype)框架,填补了这一空白,识别出了决定刺胞动物幼虫多样化形态的中尺度力学模块。通过将比较形态发生学与活性物质理论相结合,该团队证明了复杂的形态演化可以还原为一组有限的可调物理参数。
研究人员研究了包括海葵、珊瑚和水母在内的六种刺胞动物,这些物种代表了超过5亿年的演化分歧。尽管它们的成体形态各异,但这些物种都拥有一个共同的游动幼虫阶段,即浮浪幼虫(planula)。虽然所有浮浪幼虫都是轴对称的,但它们在长宽比(伸长程度)和形状极性(沿口-反口轴的不对称程度)上表现出显著的多样性。为了理解这种多样性的物理基础,作者分析了磷酸化肌球蛋白轻链(pMLC)的分布(作为收缩力的代理),并开发了一个活性表面模型,将发育中的幼虫视为一个活性粘弹性壳层。
该研究的核心发现是识别出了“力学模块”——即超细胞单元,如空间模式化的材料特性和活性力分布。研究人员发现,轴向伸长在中尺度上是一个“简单性状”,主要由单一模块驱动:外胚层和内胚层之间基底界面的肌球蛋白应力纤维的向列序(nematic order)。在基底向列序较高的物种中,如 Nematostella vectensis 和 Hydractinia symbiolongicarpus,由此产生的活性张力驱动了显著的伸长。相反,像 Cladonema pacificum 这样缺乏这种有序基底肌球蛋白的物种,则保持相对球形。
与伸长不同,形状极性表现为一个受多个模块相互作用控制的“复杂性状”。这些模块包括口部开口的几何形状、组织弯曲刚度的区域变化(通常与外胚层厚度有关)以及顶端收缩力的分布。例如,在 Nematostella 中,由于特定的口部边界条件,形状向口端变宽。在 Exaiptasia pallida(Aiptasia)等其他物种中,较厚的反口外胚层增加了局部弯曲刚度,导致反口端变宽。有趣的是,研究发现这些力学型——即力学模块的具体配置——与系统发育基本不相关,这表明塑造身体的力学策略是独立于谱系历史演化的,可能是受生态或功能压力驱动的。
研究人员通过复杂的遗传和药理干预验证了他们的模型。通过在 Nematostella 的内胚层中表达显性负性肌球蛋白,他们成功破坏了基底向列序,这特异性地消除了轴向伸长,而没有停止整体发育。更令人惊叹的是,通过抑制 JNK 信号以改变早期轴向图式,团队能够“重编程” Nematostella 的力学型。产生的幼虫表现出形状极性的反转,有效地模拟了 Aiptasia 等姐妹物种的形态。这种实验性返祖现象表明,生物体的发育库中包含潜在的形态潜力,可以通过改变力学模块的调节来激活。
这项工作代表了我们对形态演化理解的重大概念转变。通过将力学型定义为基因型和表型之间易于处理的中间体,该研究为多样化生物形态的产生提供了生物物理逻辑。其影响远超刺胞动物;力学模块框架为比较力学生物学提供了一种通用语言,有可能揭示生命树中保守的“设计原则”。此外,理性重构这些模块的能力为合成形态学和再生医学开辟了新途径,允许通过操纵组织尺度的物理力量来工程化生物形态。